



Специфичност детекције
У већини случајева, сврха дизајна прајмера је да се максимизира специфичност ПЦР-а.Ово је одређено мање или више предвидљивим утицајем многих варијабли.Једна важна варијабла је секвенца на 3′крају прајмера.
Важно је да је већа вероватноћа да ће ПЦР тестови дизајнирани за специфичност одржати високу ефикасност у широком динамичком опсегу, јер тест не производи неспецифичне производе амплификације, чиме се такмичи са ПЦР реагенсима или инхибира главну реакцију амплификације.
Наравно, у неким случајевима специфичност није најважнија, на пример, када је циљ да се квантификују блиско повезани, али различити патогени, потребни су посебни стандарди дизајна, оптимизације и верификације.
Крива топљења је стандардни метод за процену специфичности ампликона, барем у смислу да ли треба појачати једну мету.Међутим, мора се нагласити да криве топљења могу бити погрешне јер, на пример, на њих могу утицати комбиновани ефекти субоптималних прајмера и ниске концентрације шаблона.
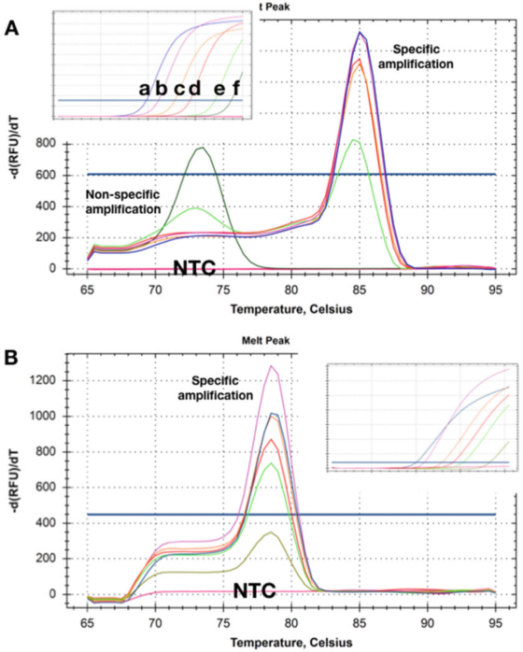
P5 |Крива топљења показује Тм помаке добијене из две детекције различитих количина две циљне ДНК.
А. При вишим концентрацијама (ад)), нема очигледног димера прајмера након што је кПЦР мерење завршено.Како се концентрација шаблона смањује на 50 копија (е), почиње да се појављује неспецифичан производ и постаје једини производ са најнижом концентрацијом (ф).
Б. Тест је забележио исти Тмс на свим циљним концентрацијама, и није било очигледног димера прајмера чак ни при најнижој концентрацији (5 копија).Када се користе ове две методе детекције, у НТЦ-има нису откривени производи појачања.
П5 приказује криве растварања добијене са узорцима у којима је шаблон присутан у различитим концентрацијама.П 5а показује да су при две најниже концентрације Тмс произведених производа неспецифичне амплификације нижи од оних специфичних ампликона.
Очигледно, овај метод детекције не може се поуздано користити за откривање циљева који постоје у ниским концентрацијама.
Занимљиво је да НТЦ, тј. узорци без ДНК уопште, нису забележили (неспецифичне) производе амплификације, што указује да позадинска геномска ДНК може да учествује у неспецифичној амплификацији/полимеризацији.
Понекад се такви основни прајмери и неспецифична амплификација не могу поправити, али је често могуће дизајнирати метод детекције који нема неспецифично појачање у било којој концентрацији шаблона и НТЦ (П 5б).
Овде, чак и снимање појачања циљне концентрације са Цк од 35 ће произвести специфичну криву растварања.Слично, НТЦ нису показивали знаке неспецифичне амплификације.Понекад, понашање детекције може зависити од матичне течности, а само неспецифична амплификација се детектује у одређеним пуферским композицијама, што може бити повезано са различитим концентрацијама Мг2+.
Стабилност детекције
Оптимизација Та је користан корак у емпиријској верификацији и процесу оптимизације кПЦР детекције.Он пружа директну индикацију робусности сета прајмера тако што показује температуру (или температурни опсег) која производи најнижи Цк без појачавања НТЦ-а.
Двострука до четворострука разлика у осетљивости можда није важна за људе са високом експресијом мРНК, али за дијагностичке тестове може значити разлику између позитивних и лажно негативних резултата.
Та својства кПЦР прајмера могу значајно да варирају.Неки тестови нису веома робусни, и ако се не изводе под оптималном Та вредношћу прајмера, брзо ће се срушити.
Ово је важно јер је ова врста детекције често проблематична у стварном свету, а чистоћа узорка, концентрација ДНК или присуство друге ДНК можда нису оптимални.
Поред тога, циљни број копија може да варира у широком опсегу, а реагенси, пластични прибор или инструменти могу да се разликују од оних који се користе приликом постављања теста.
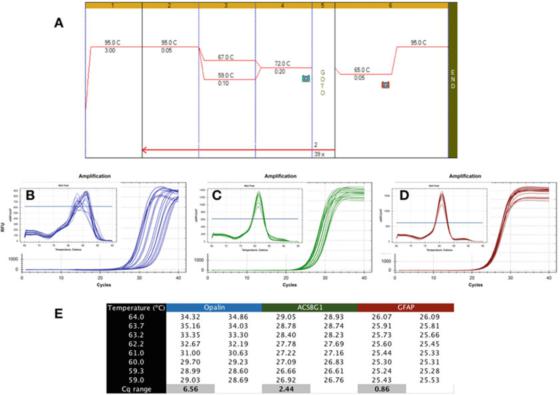
P6|Градијент температуре показује различиту робусност ПЦР детекције.
А. Користите Биолине-ов Сенсифаст СИБР мастермик (каталошки број БИО-98050) да извршите ПЦР на цДНК припремљеној од РНК људског мозга.
Б. Користите Био-Рад-ов ЦФКС кПЦР инструмент да снимите мапу амплификације и криву растварања апалена (НМ_033207, Ф: ГЦЦАТГГАГГАААГТГАЦАГАЦЦ, Р: ЦТЦАТГТГТГГГТГАТЦТЦЦТАГГ).
Ц. Графикон појачања и крива топљења АЦСБГ1 (НМ_015162.4, Ф: ЦТАЦАЦТТЦЦГГЦАЦЦАЦТГГ, Р: ГТЦЦАЦГТГАТАТТГТЦТТГАЦТЦАГ).
Д. Графикон амплификације и крива растварања ГФАП-а (НМ_002055.5, Ф: ТГГАГАГГААГАТТГАГТЦГЦТГГ, Р: ЦГААЦЦТЦЦТЦЦТЦГТГГАТЦТТЦ).
Е. Цкс забележени на различитим температурама жарења, показујући разлику у Цк забележеној под температурним градијентом од 7Ц.
П 6 показује типичан резултат непожељног теста, где је кПЦР изведен коришћењем градијента Тас између 59Ц и 67Ц (П 6а), коришћењем прајмера за три гена специфична за људски мозак.
Из графикона амплификације се може видети да су опалин прајмери далеко од идеалних јер је њихов оптимални Та опсег веома узак (слика 6б), односно, Цкс су широко распршени, што резултира значајном упоређивањем Цкс са њиховим оптималним Цкс Лов.
Овај метод детекције је нестабилан и може довести до субоптималног појачања.Стога, овај пар прајмера треба редизајнирати.Поред тога, анализа криве топљења (инсет) показује да специфичност ове методе детекције такође може бити проблематична, јер је крива топљења сваког Та различита.
Метода детекције АЦСБГ1 приказана у П 6ц је робустнија од методе детекције Опалина изнад, али је још увек далеко од идеалне и вероватно је да се може побољшати.
Међутим, наглашавамо да не постоји неопходна веза између робусности и специфичности, јер крива растварања произведена овом методом детекције показује исту вршну вредност у свим Тас (уметнутим).
С друге стране, тест робусности је много толерантнији, стварајући сличне Цкс у широком опсегу Тас, као у ГФАП тесту приказаном у П 6д.
Разлика у Цкс добијеној у истом опсегу од 8 степени Целзијуса је мања од 1, а крива растварања (уметнути) потврђује карактеристике детекције у овом температурном опсегу.Вреди напоменути да израчунати Тас и стварни Та распон могу бити веома различити.
Постоје многе смернице осмишљене да помогну истраживачима да дизајнирају ефикасне прајмере, од којих се већина заснива на давно утврђеним правилима и пуно пажње је посвећено 3′крају прајмера.Често се препоручује да се укључи Г или Ц на крају од 3' и две Г или Ц базе (ГЦ стезаљка), али не више од две од последњих 5 база.
У пракси, ова правила могу водити истраживаче, али нису нужно тачна у свим околностима.
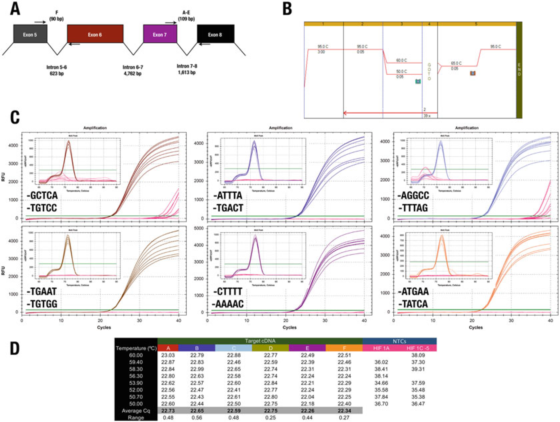
P7 |3′крај прајмера има мали утицај на специфичност или ефикасност.
А. Положај прајмера за хумани ХИФ-1α (НМ_181054.2) ген.
Б. Користите Агилент Бриллиант ИИИ СИБР Греен матичну течност (кат. бр. 600882) да појачате шест тест јединица.
Ц. Графикон амплификације и крива топљења снимљени Био-Рад-овим ЦФКС кПЦР инструментом и 3′крајним прајмерима.НТЦ су приказани црвеном бојом.
Д. Цкс запис за сваки испитни предмет
На пример, резултат у П 7 је у супротности са правилом 3′краја.Сви дизајни дају у основи исте резултате, са само две комбинације прајмера које доводе до неспецифичног појачања у НТЦ.
Међутим, не можемо подржати ефекат ГЦ клипа, јер у овом случају коришћење А или Т као максимално 30 база не смањује специфичност.
Тест Ц, где се Ф прајмер завршава на ГГЦЦ, је забележио Цкс у НТЦ, што указује да би неко можда желео да избегне ове секвенце на 30-крају.Наглашавамо да је једини начин да се одреди најбоља 3′крајна секвенца пара прајмера да се експериментално процене неки кандидати за прајмере.
Ефикасност појачања
Важно је да иако неспецифична ПЦР детекција никада не може постати специфична, ефикасност амплификације се може прилагодити и максимизирати на много различитих начина променом ензима, матичне течности, адитива и услова циклуса.
Да би се проценила ефикасност ПЦР детекције, најбоље је користити серијско разблаживање 10 или 5 пута веће од циљне нуклеинске киселине, односно „метода стандардне криве“.
Ако се ПЦР ампликони или мете синтетичке ДНК користе за генерисање стандардне криве, серијска разблажења ових мета треба да се мешају са константном количином позадинске ДНК (као што је геномска ДНК).
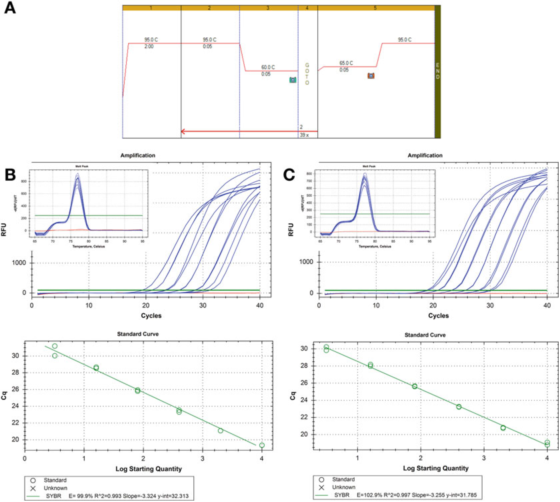
P8 |Крива разблажења за процену ефикасности ПЦР-а.
А. Користите прајмере за ХИФ-1: Ф: ААГААЦТТТТАГГЦЦГЦТЦА и Р: ТГТЦЦТГТГГТГАЦТТГТЦЦ и Агилентов Бриллиант ИИИ СИБР Греен мастермик (каталошки број 600882) за ПЦР и услове криве топљења.
Б. 100 нг РНК је реверзно транскрибовано, разблажено 2 пута, а серијски разблажени цДНК узорци су разблажени 5 пута до 1 нг хумане геномске ДНК.Крива топљења је приказана на уметку.
C. РТ реакција, разблаживање и серијско разблаживање су поновљени за други узорак цДНК, а резултати су били слични.
П 8 приказује две стандардне криве, користећи исти метод детекције на два различита узорка цДНК, резултат је иста ефикасност, око 100%, а слична је и вредност Р2, односно степен уклапања између експерименталних података и линије регресије или података Степен линеарности.
Две стандардне криве су упоредиве, али нису потпуно исте.Ако је сврха да се тачно квантификује циљ, мора се напоменути да је неприхватљиво давати прорачун броја копија без објашњења несигурности
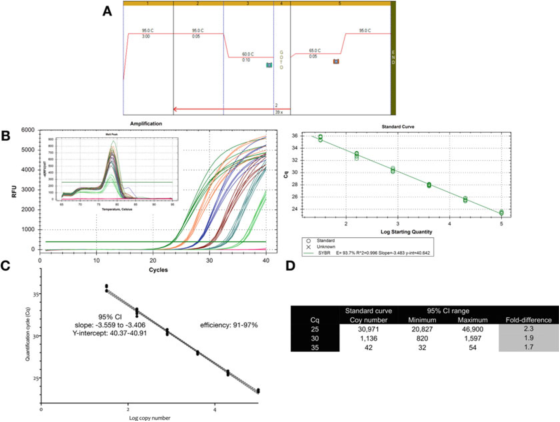
P9 |Несигурност мерења повезана са квантификацијом коришћењем стандардне криве.
А. Користите прајмере за ГАПДХ (НМ_002046) да бисте извршили ПЦР и услове криве топљења.Ф: АЦАГТТГЦЦАТГТАГАЦЦ и Р: ТААЦТГГТТГАГЦАЦАГГ и Биолине-ов Сенсифаст СИБР мастермикс (каталошки број БИО-98050).
Б. Табела појачања, крива топљења и стандардна крива снимљена Био-Рад-овим ЦФКС кПЦР инструментом.
Ц. Графикон стандардне криве и 95% интервал поверења (ЦИ).
Д. Број копија и 95% интервал поверења за три вредности Цк изведене из криве разблажења.
П 9 показује да је за оптимизовани тест инхерентна варијабилност једне стандардне криве приближно 2 пута (95% интервал поверења, минимум до максимума), што може бити најмања варијабилност која се може очекивати.
Сродних производа:
Анимал Тиссу Дирецт ПЦР комплет
Време поста: 30.09.2021



