



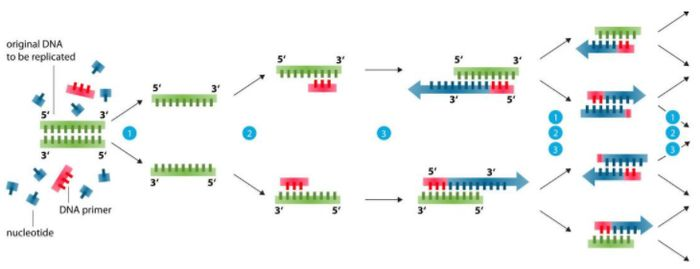
Основа дизајна прајмера (99% проблема се може решити)
1. Дужина прајмера: Уџбеник захтева 15-30бп, обично око 20бп.Стварно стање је боље да буде 18-24бп да би се осигурала специфичност, али што дужи, то бољи, предугачак прајмер ће такође смањити специфичност и смањити принос.
2. Распон амплификације прајмера: 200-500бп је прикладно, а фрагмент се може проширити на 10кб под одређеним условима.
3. Подлога прајмера: Садржај Г+Ц треба да буде 40-60%, премало Г+Ц ефекта појачања није добро, превише Г+Ц може лако да се појаве неспецифичне траке.АТГЦ се најбоље распоређује насумично, избегавајући кластере од више од 5 пуринских или пиримидинских нуклеотида.Мулти-гц за 5′ крај и међусеквенце ради повећања стабилности, избегавање богатог ГЦ на крају 3′, без ГЦ за последње 3 базе или без ГЦ за 3 од последњих 5 база.
4. Избегавајте секундарну структуру у прајмерима и избегавајте комплементацију између два прајмера, посебно комплементарност на 3' крају, у супротном ће се формирати димер прајмера и генерисати неспецифичне појачане траке.
5. Базе на 3' крају прајмера, посебно последња и претпоследња база, треба да буду стриктно упарене да би се избегао неуспех ПЦР-а због неупарених терминалних база.
6. Прајмери имају или се могу додати са одговарајућим местима цепања, а амплификована циљна секвенца би пожељно требало да има одговарајућа места цепања, што је веома корисно за анализу цепања или молекуларно клонирање.
7. Специфичност прајмера: прајмери не би требало да имају очигледну хомологију са другим секвенцама у бази података секвенци нуклеинских киселина.
8. Научите да користите софтвер: ПП5, Олиго6, ДНАстар, Вецтор НТИ, пример3 (Овај онлајн дизајн најбоље функционише).
Горњи садржај може да реши најмање 99% проблема дизајна прајмера.
Контролишите детаље дизајна прајмера
1. Дужина прајмера
Општа дужина прајмера је 18~30 база.Генерално, најважнији фактор који одређује температуру жарења прајмера је дужина прајмера.Температура жарења прајмера се генерално бира (Тм вредност -5℃), а неки директно користе Тм вредност.Следеће формуле се могу користити за грубо израчунавање температуре жарења прајмера.
Када је дужина прајмера мања од 20 бп: [4(Г+Ц)+2(А+Т)]-5℃
Када је дужина прајмера већа од 20 бп: 62,3℃+0,41℃(%ГЦ)-500/дужина-5℃
Поред тога, многи софтвери се такође могу користити за израчунавање температуре жарења, принцип израчунавања ће бити другачији, тако да понекад израчуната вредност може имати мали јаз.Да би се оптимизовале ПЦР реакције, користе се најкраћи прајмери који обезбеђују температуре жарења не мање од 54℃ за најбољу ефикасност и специфичност.
Све у свему, специфичност прајмера се повећава за фактор четири за сваки додатни нуклеотид, тако да је минимална дужина прајмера за већину примена 18 нуклеотида.Горња граница дужине прајмера није од велике важности, углавном се односи на ефикасност реакције.Због ентропије, што је прајмер дужи, то је нижа брзина којом се он жари да би се везао за циљну ДНК да би се формирао стабилан дволанчани шаблон за везивање ДНК полимеразе.
Када се користи софтвер за дизајнирање прајмера, дужина прајмера се може одредити помоћу ТМ вредности, посебно за прајмере флуоресцентног квантитативног ПЦР-а, ТМ=60℃ или тако треба контролисати.
2.ГЦ садржај
Генерално, садржај Г+Ц у секвенцама прајмера је 40%~60%, а ГЦ садржај и Тм вредност пара прајмера треба да буду координисани.Ако прајмер има озбиљну ГЦ или АТ тенденцију, одговарајућа количина А, Т или Г и Ц репа може се додати на 5' крај прајмера.
3. Температура жарења
Температура жарења треба да буде 5 ℃ нижа од температуре одвајања ланца.Ако је број база прајмера мали, температура жарења се може повећати на одговарајући начин, што може повећати специфичност ПЦР-а.Ако је број база велики, температура жарења се може на одговарајући начин смањити.Разлика у температури жарења између пара прајмера од 4℃ ~ 6℃ неће утицати на принос ПЦР-а, али у идеалном случају температура жарења пара прајмера је иста, која може да варира између 55℃ ~ 75℃.
4. Избегавајте област секундарне структуре шаблона за појачавање
Најбоље је избегавати регион секундарне структуре шаблона када бирате појачани фрагмент.Стабилна секундарна структура циљног фрагмента може се предвидети и проценити помоћу релевантног рачунарског софтвера, што је од помоћи за одабир шаблона.Експериментални резултати показују да је експанзија често неуспешна када је слободна енергија (△Г) региона који се шири мања од 58,6 лкЈ/мол.
5. Неусклађеност са циљном ДНК
Када је амплификована циљна ДНК секвенца велика, прајмер се може везати за више делова циљне ДНК, што резултира појавом више трака у резултату.Овај пут је потребно користити БЛАСТ тестирање софтвера, веб страницу:хттп://ввв.нцби.нлм.них.гов/БЛАСТ/.Изаберите Поравнај две секвенце (бл2сек).
Лепљење секвенци прајмера у зону 1 и секвенци циљне ДНК у зону 2 је заменљиво, а БЛАСТ израчунава комплементарне, антисенс и друге могућности, тако да корисници не морају да примећују да ли су оба ланца чулни ланци.Такође можете да унесете ГИ број ако знате ГИ број секвенце у бази података, тако да не морате да лепите велики део низа.На крају, кликните на Поравнај на 3 да видите да ли прајмер има више хомологних места у циљној ДНК.
6. Терминал за прајмер
Крај 3' прајмера је место где почиње продужетак, тако да је важно спречити да неслагања почну тамо.Крај 3' не би требало да буде више од 3 узастопна Г или Ц, јер ће то проузроковати да се прајмер грешком покрене у региону секвенце обогаћивања Г+Ц.3' крај не може да формира никакву секундарну структуру, осим у специјалним ПЦР (АС-ПЦР) реакцијама, 3' крај прајмера не може бити неусклађен.На пример, ако је кодирајући регион појачан, 3' крај прајмера не би требало да буде прекинут на трећој позицији кодона, јер је трећа позиција кодона склона дегенерацији, што ће утицати на специфичност и ефикасност амплификације.Када користите прајмере за анексију, погледајте табелу употребе кодона, обратите пажњу на биолошке преференције, немојте користити анексијске прајмере на крају 3′ и користите већу концентрацију прајмера (1уМ-3уМ).
7. Секундарна структура прајмера
Сами прајмери не би требало да имају комплементарне секвенце, иначе ће се сами прајмери савијати у структуре укосница, а ова секундарна структура ће утицати на везивање прајмера и шаблона због стеричне сметње.Ако се користи вештачко расуђивање, континуиране комплементарне базе самих прајмера не би требало да буду веће од 3бп.Не би требало да постоји комплементарност између два прајмера, посебно треба избегавати комплементарно преклапање 3' краја да би се спречило стварање димера прајмера.Генерално, не би требало да постоји више од 4 узастопне хомологије или комплементарности између пара прајмера.
8. Додајте маркере или локусе
5' крај има мали утицај на специфичност амплификације и стога се може модификовати без утицаја на специфичност амплификације.Модификација 5' краја прајмера је укључивала: додавање места за рестрикцију ензима;Означени биотин, флуоресценција, дигоксин, Еу3+, итд. Увести секвенце ДНК које везују протеине;Увођење места мутације, уметање и изостанак секвенци мутације и увођење промоторских секвенци, итд. Додатне базе ће мање-више утицати на ефикасност амплификације и повећати шансу за формирање димера прајмера, али се морају направити неки уступци за следећи корак.Додатне секвенце које не постоје на циљној секвенци, као што су рестрикциона места и секвенце промотера, могу се додати на 5′ крај прајмера без утицаја на специфичност.Ове секвенце нису укључене у израчунавање вредности прајмера Тм, али их треба тестирати на комплементарност и унутрашњу секундарну структуру.
9. Субцлонес
Већину времена, ПЦР је само прелиминарно клонирање, а онда треба да субклонирамо циљни фрагмент у различите векторе, тако да треба да дизајнирамо додатне базе за следећу операцију у ПЦР кораку.
Неке секвенце дизајниране за субклонирање су сажете у наставку.
Додато је рестрикционо место ендонуклеазе
Додавање рестрикцијских места ензима је најчешће коришћена метода за субклонирање ПЦР производа.Генерално, место цепања је шест база, поред 5 'краја места цепања потребно је додати 2 ~ 3 заштитне базе.Међутим, број заштитних база које захтевају различити ензими је различит.На пример, СалⅠ не захтева заштитну подлогу, ЕцоРⅤ захтева 1 заштитну базу, НотⅠ захтева 2 заштитне базе, а Хинд Ⅲ захтева 3 заштитне базе.
ЛИЦ додаје реп
Пуни назив ЛИЦ-а је клонирање независно од лигације, метод клонирања који је изумео Навоген посебно за свој део пЕТ вектора.ПЕТ носач припремљен ЛИЦ методом има некомплементарне 12-15 базних једноланчаних лепљивих крајева, који допуњују одговарајуће лепљиве крајеве на фрагменту циљног уметања.За потребе амплификације, секвенца прајмера 5′ уметнутог фрагмента треба да допуни ЛИЦ вектор.Екстранектна активност 3′→5′ Т4 ДНК полимеразе може да формира један ланац лепљивог краја на уметнутом фрагменту након кратког времена.Пошто се производ може формирати само међусобним жарењем припремљеног фрагмента инсерта и вектора, овај метод је веома брз и ефикасан, а ради се о усмереном клонирању.
Усмерени ТА клон додати реп
ТА клонирање није било у стању да усмери фрагмент у вектор, па је касније Инвитроген увео вектор који је могао да циља клонирање, који је садржао четири истакнуте базе ГТГГС на једном крају.Стога, у дизајну ПЦР прајмера треба додати комплементарне секвенце у складу са тим, тако да се фрагменти могу „оријентисати“.
Ако немате довољно времена, можете пробати директну синтезу, комбинујући ген са вектором, што ми код мускулариста називамо синтезом ЕТ гена.
Д. Метода клонирања у фузији
Није потребна лигаза, није потребна дуга реакција.Све док се секвенца на оба краја линеаризованог вектора уводи у дизајн прајмера, онда се ПЦР производ и линеаризовани вектор додају у инфузиони раствор ензима који садржи БСА и стављају на собну температуру пола сата, трансформација се може извршити.Овај метод је посебно погодан за конверзију великог обима.
10. Спајање прајмера
Понекад су о дизајну прајмера познате само ограничене информације о секвенци.На пример, ако је позната само секвенца аминокиселина, може се дизајнирати прајмер за спајање.Прајмер спајања је мешавина различитих секвенци које представљају све различите могућности база које кодирају једну аминокиселину.Да бисте повећали специфичност, можете погледати табелу употребе кодона да бисте смањили анексију у складу са преференцијама основне употребе различитих организама.Хипоксантин се може упарити са свим базама да би се смањила температура жарења прајмера.Немојте користити приложене базе на 3′ крају прајмера јер је жарење последње 3 базе на 3′ крају довољно да започне ПЦР на погрешном месту.Користе се веће концентрације прајмера (1 μМ до 3 μМ) јер прајмери у многим смешама за анексију нису специфични за циљни шаблон.
ПЦР сировинеконтролу
1. Количина прајмера
Концентрација сваког прајмера је 0,1 ~ 1 умол или 10 ~ 100 пмол.Боље је постићи жељени резултат са најмањом количином прајмера.Висока концентрација прајмера ће изазвати неусклађеност и неспецифично појачање, и повећати шансу за формирање димера између прајмера.
2. Концентрација прајмера
Концентрација прајмера утиче на специфичност.Оптимална концентрација прајмера је углавном између 0,1 и 0,5 μМ.Веће концентрације прајмера доводе до амплификације неспецифичних производа.
3. Температура жарења прајмера
Други важан параметар за прајмере је температура топљења (Тм).Ово је температура када је 50% прајмера и комплементарних секвенци представљено као дволанчани ДНК молекули.Тм је потребан за подешавање температуре жарења ПЦР-а.У идеалном случају, температура жарења је довољно ниска да обезбеди ефикасно жарење прајмера са циљном секвенцом, али довољно висока да смањи неспецифично везивање.Разумна температура жарења од 55℃ до 70℃.Температура жарења је генерално подешена за 5 ℃ нижа од Тм прајмера.
Постоји неколико формула за подешавање Тм, које се веома разликују у зависности од формуле која се користи и редоследа прајмера.Пошто већина формула даје процењену вредност Тм, све температуре жарења су само почетна тачка.Специфичност се може побољшати анализом неколико реакција које прогресивно подижу температуру жарења.Почните испод процењене Тм-5℃, и постепено повећавајте температуру жарења у порасту од 2℃.Виша температура жарења ће смањити формирање димера прајмера и неспецифичних производа.За најбоље резултате, два прајмера треба да имају приближне вредности Тм.Ако је разлика Тм парова прајмера већа од 5 ℃, прајмери ће показати значајан погрешан почетак коришћењем ниже температуре жарења у циклусу.Ако су два прајмера Тм различита, подесите температуру жарења на 5℃ нижу од најниже Тм.Алтернативно, да би се повећала специфичност, пет циклуса се може извести прво на температурама жарења које су дизајниране за више Тм, након чега следе преостали циклуси на температурама жарења дизајнираним за ниже Тм.Ово омогућава добијање делимичне копије одредишног шаблона у тешким условима.
4. Чистоћа и стабилност прајмера
Стандардна чистоћа прилагођених прајмера је адекватна за већину ПЦР апликација.Уклањање бензоил и изобутилил група одсољавањем је минимално и стога не омета ПЦР.Неке апликације захтевају пречишћавање да би се уклониле све секвенце које нису пуне дужине у процесу синтезе.Ове скраћене секвенце настају зато што ефикасност хемије синтезе ДНК није 100%.Ово је кружни процес који користи поновљене хемијске реакције док се свака база додаје да би се направила ДНК од 3′ до 5′.Можете пропасти ни у једном циклусу.Дужи прајмери, посебно они већи од 50 база, имају велики удео скраћених секвенци и могу захтевати пречишћавање.
На принос прајмера утиче ефикасност синтетичке хемије и методе пречишћавања.Биофармацеутске компаније, као што су Цитологи и Схенгонг, све користе минималну ОД јединицу како би осигурале укупан излаз олигонуклеозида.Прилагођени прајмери се испоручују у облику сувог праха.Најбоље је поново растворити прајмере у ТЕ тако да коначна концентрација буде 100 μМ.ТЕ је бољи од дејонизоване воде јер је пХ воде често кисел и изазива хидролизу олигонуклеозида.
Стабилност прајмера зависи од услова складиштења.Суви прах и растворени прајмери треба да се чувају на -20℃.Прајмери растворени у ТЕ у концентрацијама већим од 10 μМ могу се стабилно чувати на -20℃ током 6 месеци, али се могу чувати само на собној температури (15℃ до 30℃) мање од 1 недеље.Прајмери са сувим прахом могу се чувати на -20 Ц најмање 1 годину и на собној температури (15 Ц до 30 Ц) до 2 месеца.
5. Ензими и њихове концентрације
Тренутно, коришћена Так ДНК полимераза је у основи ензим генског инжењеринга који синтетише колиформне бактерије.Количина ензима потребна за катализацију типичне ПЦР реакције је око 2,5У (односи се на укупну запремину реакције од 100ул).Ако је концентрација превисока, то може довести до неспецифичног појачања;ако је концентрација прениска, количина синтетичког производа ће се смањити.
6. Квалитет и концентрација дНТП
Квалитет дНТП је уско повезан са концентрацијом и ефикасношћу ПЦР амплификације.ДНТП прах је зрнаст, а његова варијабилност губи своју биолошку активност ако се неправилно складишти.Раствор дНТП је кисео и треба га користити у високој концентрацији, са 1М НаОХ или 1М Трис.ХЦЛ пуферским раствором да би се подесио пХ на 7,0 ~ 7,5, мала количина под-паковања, замрзнуто складиштење на -20℃.Вишеструко замрзавање-одмрзавање ће разградити дНТП.У ПЦР реакцији, дНТП би требало да буде 50 ~ 200 умол/Л.Посебно треба обратити пажњу на то да концентрација четири ДНТПС треба да буде једнака (једнака припрема мола).Ако је концентрација било ког од њих различита од осталих (виша или нижа), доћи ће до неусклађености.Прениска концентрација ће смањити принос ПЦР производа.дНТП се може комбиновати са Мг2+ и смањити концентрацију слободног Мг2+.
7. Шаблон (циљни ген) нуклеинска киселина
Количина и степен пречишћавања шаблонске нуклеинске киселине је једна од кључних карика за успех или неуспех ПЦР-а.Традиционалне методе пречишћавања ДНК обично користе СДС и протеазу К за варење и одлагање узорака.Главне функције СДС-а су: растварање липида и протеина на ћелијској мембрани, уништавајући на тај начин ћелијску мембрану растварањем мембранских протеина, и дисоциацију нуклеарних протеина у ћелији, СДС се такође може комбиновати са протеинима и таложити;Протеаза К може да хидролизује и вари протеине, посебно хистоне везане за ДНК, а затим користи органски растварач фенол и хлороформ за екстракцију протеина и других ћелијских компоненти и користи етанол или изопропил алкохол за таложење нуклеинске киселине.Екстрахована нуклеинска киселина се може користити као шаблон за ПЦР реакције.За узорке за општу клиничку детекцију, брза и једноставна метода се може користити за растварање ћелија, лизат патогена, варење и уклањање протеина из хромозома до слободних циљних гена, и директно коришћење за ПЦР амплификацију.Екстракција шаблона РНК обично користи методу гванидин изотиоцијаната или протеазе К како би се спречила разградња РНК РНА.
8. Концентрација Мг2+
Мг2+ има значајан утицај на специфичност и принос ПЦР амплификације.У општој ПЦР реакцији, када је концентрација различитих дНТП 200 умол/Л, одговарајућа концентрација Мг2+ је 1,5 ~ 2,0 ммол/Л.Концентрација Мг2+ је превисока, специфичност реакције се смањује, долази до неспецифичне амплификације, прениска концентрација ће смањити активност Так ДНК полимеразе, што резултира смањењем продуката реакције.
Јони магнезијума утичу на неколико аспеката ПЦР-а, као што је активност ДНК полимеразе, што утиче на принос;Други пример је жарење прајмера, што утиче на специфичност.дНТП и шаблон се везују за јон магнезијума, смањујући количину слободног јона магнезијума потребног за активност ензима.Оптимална концентрација јона магнезијума варира за различите парове прајмера и шаблоне, али типична почетна концентрација за ПЦР са 200 μМ дНТП је 1,5 мМ (напомена: За квантитативни ПЦР у реалном времену, користите раствор магнезијумових јона од 3 до 5 мМ са флуоресцентном сондом).Веће концентрације слободних јона магнезијума повећавају принос, али и повећавају неспецифично појачање и смањују верност.Да би се одредила оптимална концентрација, титрације јона магнезијума су вршене у корацима од 0,5 мМ од 1 мМ до 3 мМ.Да би се смањила зависност од оптимизације јона магнезијума, може се користити Платинум Так ДНК полимераза.Платинум Так ДНК полимераза је у стању да одржи функцију у ширем опсегу концентрација магнезијумових јона од Так ДНК полимеразе и стога захтева мање оптимизације.
9. Адитиви који промовишу Пцр
Оптимизација температуре жарења, дизајна прајмера и концентрације јона магнезијума довољна је за високо специфично појачање већине шаблона;међутим, неки шаблони, укључујући и оне са високим садржајем ГЦ, захтевају додатне мере.Адитиви који утичу на температуру топљења ДНК пружају још један начин за побољшање специфичности производа и приноса.За најбоље резултате потребна је потпуна денатурација шаблона.
Поред тога, секундарна структура спречава везивање прајмера и проширење ензима.
ПЦР адитиви, укључујући формамид, ДМСО, глицерин, бетаин и ПЦРк Енханцер Солутион, побољшавају амплификацију.Њихов могући механизам је смањење температуре топљења, чиме се помаже жарење прајмера и помаже проширење ДНК полимеразе кроз регион секундарне структуре.ПЦРк решење има и друге предности.Потребна је минимална оптимизација јона магнезијума када се користи са Платинум Так ДНК полимеразом и Платинум Пфк ДНК полимеразом.Дакле, Платинум техника је комбинована са додатком за повећање специфичности уз истовремено смањење зависности трећег приступа, оптимизације јона магнезијума.За најбоље резултате треба оптимизовати концентрацију адитива, посебно ДМСО, формамида и глицерола, који инхибирају Так ДНК полимеразу.

10. Хот старт
Хот старт ПЦР је једна од најважнијих метода за побољшање специфичности ПЦР-а поред доброг дизајна прајмера.Иако је оптимална температура елонгације Так ДНК полимеразе 72℃, полимераза остаје активна на собној температури.Дакле, неспецифични производи се производе када је температура држања нижа од температуре жарења током припреме ПЦР реакције и на почетку термичког циклуса.Једном формирани, ови неспецифични производи се ефикасно појачавају.Хот-старт ПЦР је посебно ефикасан када су места која се користе за дизајн прајмера ограничена локацијом генетских елемената, као што су мутације усмерене на место, клонирање експресије или конструкција и манипулација генетским елементима који се користе за ДНК инжењеринг.
Уобичајени метод за ограничавање активности Так ДНК полимеразе је припремање ПЦР реакционог раствора на леду и постављање у претходно загрејани ПЦР апарат.Ова метода је једноставна и јефтина, али не довршава активност ензима и стога не елиминише у потпуности амплификацију неспецифичних производа.
Термални прајминг одлаже синтезу ДНК инхибирајући есенцијалну компоненту све док ПЦР апарат не достигне температуру денатурације.Већина метода ручног термичког иницирања, укључујући одложено додавање Так ДНК полимеразе, су гломазне, посебно за апликације велике пропусности.Друге методе термичког прајминга користе штит од воска за затварање есенцијалне компоненте, укључујући јоне магнезијума или ензиме, или да физички изолују реактивне компоненте, као што су шаблони и пуфери.Током термичког циклуса, различите компоненте се ослобађају и мешају заједно док се восак топи.Као и метода ручног врућег покретања, метода заштите од воска је гломазна и склона контаминацији и није погодна за апликације велике пропусности.
Платинум ДНК полимераза је погодна и ефикасна за ПЦР са аутоматским стартом.Платинум Так ДНК полимераза се састоји од рекомбинантне Так ДНК полимеразе комбиноване са моноклонским антителом против Так ДНК полимеразе.Антитела су формулисана ПЦР-ом да инхибирају активност ензима током дужег одржавања температуре.Так ДНК полимераза је пуштена у реакцију током 94℃ изолације корака денатурације, враћајући пуну активност полимеразе.За разлику од хемијски модификоване Так ДНК полимеразе за термичку иницијацију, платинасти ензим не захтева продужену изолацију на 94℃ (10 до 15 минута) да би активирао полимеразу.Са ПлатинумТак ДНК полимеразом, 90% активности Так ДНК полимеразе је обновљено након 2 минута на 94 ℃.

11. Нест-ПЦР
Узастопни кругови амплификације коришћењем угнежђених прајмера могу побољшати специфичност и осетљивост.Први круг је стандардно појачање од 15 до 20 циклуса.Мала фракција иницијалног производа амплификације је разблажена 100 до 1000 пута и додата у други круг амплификације током 15 до 20 циклуса.Алтернативно, почетни амплификовани производ се може одредити пречишћавањем гела.У другом кругу амплификације се користи угнежђени прајмер, који се може везати за циљну секвенцу унутар првог прајмера.Употреба угнежђеног ПЦР-а смањује могућност амплификације вишеструких циљних места јер постоји неколико циљних секвенци комплементарних оба сета прајмера.Исти укупан број циклуса (30 до 40) са истим прајмерима је појачао неспецифична места.Нестед ПЦР повећава осетљивост ограничених циљних секвенци (нпр. ретке мрнас) и побољшава специфичност тешког ПЦРС (нпр. 5′ РАЦЕ).
12. Силазни ПЦР
Опадајући ПЦР побољшава специфичност коришћењем чврстих услова жарења за првих неколико циклуса ПЦР-а.Циклус почиње при температури жарења приближно 5℃ вишој од процењене Тм, а затим се сваки циклус смањује за 1℃ до 2℃ све док температура жарења не падне испод Тм 5℃.Само одредишни шаблон са највећом хомологијом ће бити појачан.Ови производи настављају да се шире у наредним циклусима, истискујући појачане неспецифичне производе.Десцендентни ПЦР је користан за методе где степен хомологије између прајмера и циљног шаблона није познат, као што је АФЛП ДНК отисак прста.
Повезани ПЦР комплети
2× ПЦР херојTMМик систем има већу толеранцију на ПЦР инхибиторе од обичног ПЦР Мик система и лако се носи са ПЦР амплификацијом различитих сложених шаблона.Јединствени реакциони систем и високоефикасни Так Херо чине да ПЦР реакција има већу ефикасност амплификације, специфичност и осетљивост.
Већа ефикасност појачања
Има активност 5'→3' ДНК полимеразе и 5'→3' егзонуклеазне активности, без активности 3'→5' егзонуклеазе.
Реал Тиме ПЦР Еасиᵀᴹ-СИБР Греен И Кит
Специфичан—оптимизовани пуфер и Так ензим са врућим стартом могу спречити неспецифично појачавање и формирање димера прајмера
Висока осетљивост—може открити мале копије шаблона
Комплет користи јединствени Форегене реагенс реверзне транскрипције и Форегене ХотСтар Так ДНК полимеразу у комбинацији са јединственим реакционим системом за ефикасно побољшање ефикасности амплификације и специфичности реакције.
Време поста: 09.05.2023



